Structure cristallographique d’une partie du système de sécrétion de type VI résolue sur PROXIMA-1
Parmi les multiples systèmes utilisés par les bactéries pour survivre et se multiplier, le « T6SS », système de sécrétion de type VI, est l’un des principaux chez les bactéries dites Gram négatif, qui incluent de nombreuses espèces potentiellement pathogènes pour l’Homme (ex : E. coli, P. aeruginosa). T6SS est un complexe formé de plus de 10 protéines différentes qui permet à la bactérie de cibler et injecter des toxines dans ses cellules « proies » (procaryotes ou eucaryotes), après avoir perforé leur enveloppe. Afin de comprendre avec précision les mécanismes moléculaires qui régissent l’activité de sécrétion de ce complexe, des chercheurs de l’Institut Pasteur et du CNRS ont étudié la structure de chacun des éléments qui le composent, notamment sur PROXIMA-1 et PROXIMA-2A. Leurs résultats ont été publiés dans Nature.
Les bactéries Gram négatif possèdent une double membrane, interne/externe, séparées par un espace appelé le périplasme. Le système de sécrétion TSS6 est ancré dans la bactérie comme une seringue traversant cette double membrane : le côté « piston » est à l’intérieur, dans le cytoplasme, et l’extrémité peut pointer vers l’extérieur pour viser les cibles – mais dans ce cas, contrairement à une seringue, l’extrémité portant les toxines est éjectée. C’est pourquoi les chercheurs comparent aussi TSS6 à une nano-arbalète envoyant ses flèches sur les cibles de la bactérie. La partie transmembranaire qui permet l’ancrage du complexe dans la bactérie est formée de trois protéines, TssL et TssM positionnées dans la membrane interne, et TssJ, une protéine portant un groupement lipidique, située dans la membrane externe.
Dans le but de connaître précisément la structure des différents composants de ce complexe, les équipes de MM. Cambillau, Cascales et Fronzes ont réalisé des expériences de biocristallographie sur les lignesPROXIMA-1 et PROXIMA-2A de SOLEIL. Une partie de la structure de la protéine TssM a ainsi été résolue sur PROXIMA-1.
Les protéines transmembranaires sont difficiles à cristalliser ; les chercheurs ont travaillé sur un long fragment de TssM obtenu par protéolyse contrôlée (TssM32Ct; acides aminés 836–1129). Pour faciliter l’étape de résolution de structure, un « nanobodie » (anticorps à domaine unique) a été lié à TssM32Ct et c’est l’ensemble qui a été cristallisé. Ensuite, la résolution de structure s’est effectuée par remplacement moléculaire en utilisant la structure 3D du nanobodie nb25 fixé à la protéine étudiée (cf fig. A).
Ces données, associées à une structure résolue à l’ESRF (sur un autre fragment de TssM, appelé TssM26Ct, lié à la protéine TssJ), ainsi qu’à des résultats de microscopie électronique en coloration négative, ont permis de comprendre que la partie transmembranaire TssJLM possède une symétrie d’ordre 5, et de proposer un modèle d’assemblage des différentes protéines impliquées. Ce modèle consisterait en 5 dimères de chaque protéine, qui s’assemblent pour former un complexe traversant la membrane interne, le périplasme, et est ancré dans la membrane externe via la partie lipidique de TssJ (cf figure C). Les scientifiques pensent que l’assemblage d’autres sous-unités de T6SS avec cette partie transmembranaire doit induire un changement dans l’une des extrémités de TssM, qui traverse alors la membrane externe.
Les autres protéines composant T6SS constitueraient alors la « flèche » et le système de propulsion de la nano-arbalète.
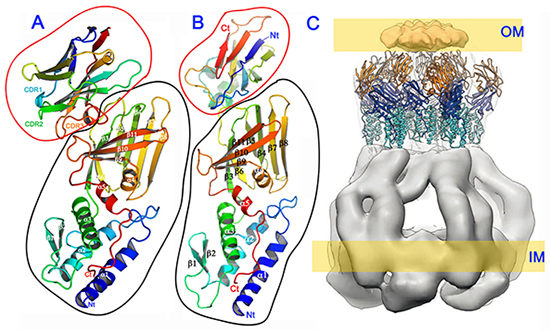
A) Représentation schématique en rubans de la structure du complexe TssM32Ct–nb25 (données de diffraction collectées sur PROXIMA1) : le fragment TssM26Ct est entouré en noir, et le nanobodie nb25 entouré en rouge. B) Représentation schématique en rubans de la structure du complexe TssM26Ct–TssJ (données collectées à l’ESRF, ligne ID23-1) : le fragment TssM26Ct est entouré en noir, et TssJ entouré en rouge. C) Modèle des 10 copies du complexe TssM26Ct–TssJ (rubans) positionné à l’intérieur du volume déterminé par microscopie électronique, qui correspond à TssJ et aux domaines périplasmiques 3 et 4 de TssM extraits à la fois des piliers internes et externes de la pointe de T6SS.