Les canaux ioniques pentamériques (5 sous-unités) sensibles à un ligand, ou pLGIC, constituent une vaste famille de récepteurs ionotropes, omniprésents dans le règne animal. Chez les vertébrés, cette famille comprend d'une part les récepteurs de la sérotonine et les récepteurs de l'acétylcholine de type nicotiniques (à perméabilité cationique), et d'autre part les récepteurs GABA et ceux de la glycine (à perméabilité anionique). Lire également l’actualité http://www.synchrotron-soleil.fr/Soleil/ToutesActualites/2013/PX1-ELIC-PNAS.
Chez les pLGIC, la liaison d’un agoniste2 à leur domaine extracellulaire provoque l'ouverture de leur domaine transmembranaire, constitué de quatre hélices α (nommées M1 à M4). Tous ces récepteurs partagent une architecture moléculaire conservée, basée sur une symétrie d'ordre cinq, avec les ions traversant un pore transmembranaire central qui est entouré par les hélices M2.
Les mécanismes moléculaires à l'origine de la perméation et de la sélectivité de charge des ions sont peu connus (Fig. 1). La structure cristalline de GLIC, un homologue bactérien de cette famille, a été élucidée à une résolution de 2,9 Å avec une forme ouverte du canal. Il s'agit d'une avancée significative pour l'étude de la perméation ionique dans les pLGIC. Le pore de GLIC est entouré par les hélices M2 qui présentent successivement six anneaux d’acides aminés dans la lumière du canal, avec de haut en bas un anneau de Glu243, trois anneaux de résidus hydrophobes Ile240, Ala237 et Ile233, deux anneaux de résidus polaires Ser230 et Thr226, et enfin Glu222. Le pore a une forme d'entonnoir, avec un grand diamètre supérieur de 11 Å et un diamètre du goulot d'étranglement de 5 Å aux niveaux 226 et 222, qui abritent le filtre de sélectivité de charge.
Combiner la cristallographie aux rayons X, l'électrophysiologie, les simulations de dynamique moléculaire et les calculs électrostatiques permet d’obtenir de nouvelles informations pertinentes sur les mécanismes moléculaires de la perméation ionique dans les pLGIC. Ces mécanismes sont complexes car ils mettent en jeu certains acides aminés des protéines, des ions, et des molécules d'eau qui interagissent de manière dynamique et transitoire pendant le passage des ions au travers du canal protéique.
L'hydratation dans le pore d'un canal ionique révélée avec un niveau de détails inégalé
La compréhension des mécanismes moléculaires de la perméation ionique dans les pLGIC exige de documenter les interactions entre les ions, les molécules d'eau et les chaînes latérales des acides aminés des M2 entourant le pore, acides aminés supposés jouer un rôle clé dans le transport ionique. Une telle description est difficile à obtenir à partir des structures disponibles de pLGIC, en raison de la faible résolution des structures obtenues précédemment par cristallographie aux rayons X.
Pour comprendre le mécanisme moléculaire de la perméation ionique dans les pLGIC, des chercheurs de l’Institut Pasteur et de l’Institut de Biologie et Chimie des Protéines de Paris ont résolu la structure de GLIC à 2,4 Å, la résolution la plus élevée jamais obtenue sur cette famille de protéines. Ces données, collectées sur la ligne PROXIMA1, révèlent pour la première fois la géométrie d'hydratation dans le pore d'un pLGIC (Fig. 2).
On observe ainsi des molécules d'eau ordonnées au niveau de deux anneaux d’acides aminés hydroxylés (Ser 230 et Thr 226) qui contribuent au filtre de sélectivité ionique. Deux pentagones d'eau sont observés, l'un pseudo-cristallin auto-stabilisé et l'autre plus grand, avec un ion sodium entre les deux. La présence d'un ion Na+ à cet emplacement a été confirmée en substituant le Na+ avec des cations possédant des propriétés de dispersion anomales (voir ci-dessous). Le deuxième pentagone est directement impliqué dans la couche d'hydratation de l'ion Na+. Ces molécules d'eau interagissent à la fois avec l'ion Na+ et avec l'atome d'oxygène du groupe carbonyle de la chaîne principale de Glu-2’.
Cartographie des sites de liaison des molécules de solvant
Pour caractériser entièrement les implications fonctionnelles des molécules d'eau organisées observées dans le pore ainsi que leur rôle dans la perméation ionique, il est nécessaire d'identifier les sites de liaison d'ions monovalents dans la structure de GLIC. Pour cela, la protéine a été co-cristallisée avec Br-, Cs+, ou Rb+. Des données aux rayons X ont été collectées à des longueurs d'onde optimisées pour chacun de ces dérivés afin de maximiser le signal anomal (Fig. 3). Ces expériences ont été réalisées sur la ligne PROXIMA1 afin de bénéficier du fait que la longueur d'onde des rayons X y est accordable. Cette technique a permis d’identifier des sites de liaison ionique. Ils sont principalement regroupés dans deux régions distinctes : dans le domaine extracellulaire (DEC) autour du bord du vestibule du canal, et dans le domaine transmembranaire près de l'extrémité cytoplasmique du pore. Les sites de liaison ionique identifiés dans le DEC sont importants pour comprendre le processus de déclenchement de l'ouverture et de la fermeture, tandis que les sites de liaison ionique identifiés dans le pore aident à la compréhension du mécanisme de perméation.
Globalement, ces résultats suggèrent que les ions en perméation occupent au moins deux positions privilégiées dans la région la plus étroite du pore, dont il a été montré qu’elle joue le rôle de filtre de sélectivité pour ces canaux.
Vers un mécanisme de perméation des ions dans les canaux ioniques sensibles à un ligand
Pour continuer à explorer les rôles de ces molécules d'eau et de ces sites de liaison ionique, les acides aminés tapissant le pore ont été remplacés par mutation et testés par électrophysiologie sur canal unique. Des simulations et des calculs électrostatiques ont complété la description de l'hydratation dans le pore. En particulier, les simulations d'un cation passant au travers du pore révèlent que les pentagones d'eau observés dans le cristal contribuent activement à la translocation pendant le passage de l'ion au travers du pore. Ces molécules d'eau sont importantes pour le franchissement des barrières d'étranglement hydrophobes. En raison de la forte conservation des séquences parmi les acides aminés des hélices M2 tapissant le pore dans la famille pLGIG, les conclusions présentées dans cette étude pourraient être transposées à l'ensemble des canaux pLGIC (Fig. 4).

Figure 1 : Structure du canal ionique GLIC. Représentations stylisées de la structure de GLIC vue de côté (seules deux sous-unités sont représentées pour des raisons de clarté) et du domaine transmembranaire (DTM) vu du côté extracellulaire. Le DTM forme un canal étroit tapissé par les hélices M2 des cinq sous-unités.
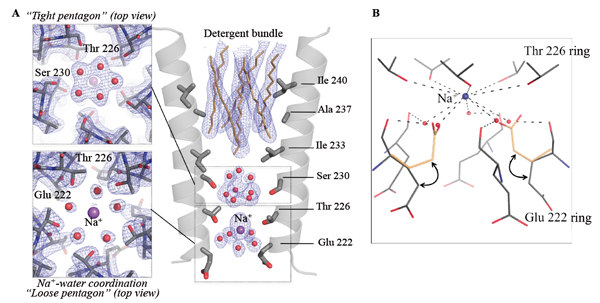
Figure 2 : Molécules d'eau structurellement ordonnées dans le pore de GLIC. (A) Vue agrandie du pore avec les hélices M2 en représentation stylisée, et les chaînes latérales des acides aminés tapissant le pore en représentation en bâtons. Un « pentagone resserré » de molécules d'eau, auto-stabilisé et bien résolu, est présent au niveau de l’acide aminé Ser 230 (en haut à gauche). Un atome Na+ et cinq molécules d'eau liées sont observés à l'extrémité intracellulaire du pore (en bas à gauche). Les détergents sont représentés par des bâtons tandis que l'eau et les atomes Na+ sont représentés par des sphères. (B) Réseau d'interaction entre les molécules d'eau, les ions Na+ et les résidus M2 tapissant le pore. La conformation alternative adoptée par Glu 226 est également représentée (en jaune).
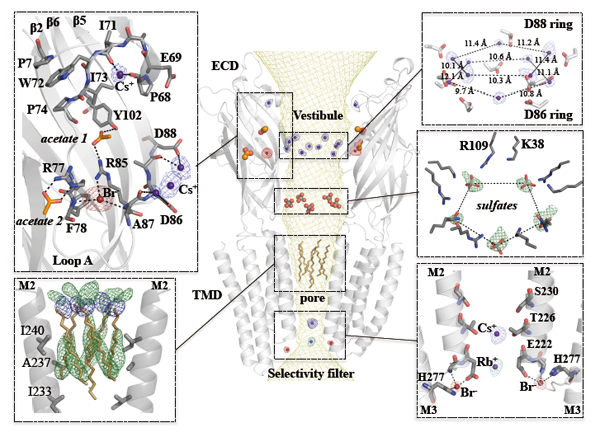
Figure 3 : Sites de liaison pour cation, anion et détergent dans le pore de GLIC. Vue centrale : Représentation stylisée de la structure de GLIC vue de côté avec la surface du canal accessible au solvant représentée par un maillage (en jaune). Les sous-figures du haut et du bas représentent les vues agrandies du vestibule du canal ainsi que du pore transmembranaire présentant les interactions entre la protéine et les molécules de solvant. (Sous-figure inférieure gauche) Orientation du détergent dodécyl-β-D-séléno-maltoside (Se-DDM). (En bas à droite) Sites de liaison ionique sur le filtre de sélectivité. Les cartes anormales calculées pour Br- (en rouge), Cs+ (en bleu) et Rb+ (en cyan) sont représentées en superposition sous forme de maillage avec les ions associés représentés par des sphères.
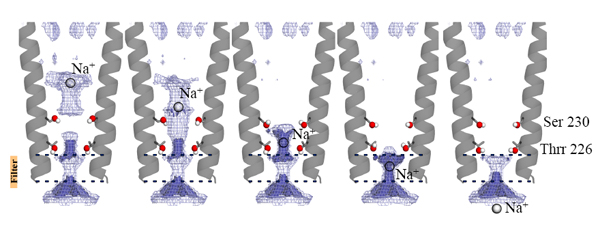
Figure 4 : Vers un mécanisme de perméation des ions dans les pLGIC. Jeu optimisé de configurations des chaînes latérales de Ser 230 et Thr 226 maximisant la densité électronique de perméation à cinq altitudes dans le pore (indiquées par une sphère blanche sur l'axe du pore), de l'extrémité extracellulaire à intracellulaire (de gauche à droite). Les groupes OH des résidus 230 et 226 sont représentés par des sphères. La région favorable pour l'ion en perméation est représentée par un maillage bleu. Les altitudes des ions observés expérimentalement dans GLIC sont indiquées par des lignes pointillées.
1 - Récepteur ionotrope : protéine membranaire interagissant avec un ligand, dont la fixation provoque l’ouverture d’un canal dans la protéine. Ce canal permet alors la traversée de la membrane par des ions.
2 – Agoniste : se dit d’un ligand dont la fixation à un récepteur provoque un changement de conformation de ce dernier, ce qui induit une réponse moléculaire (ici : traversée du canal par les ions).